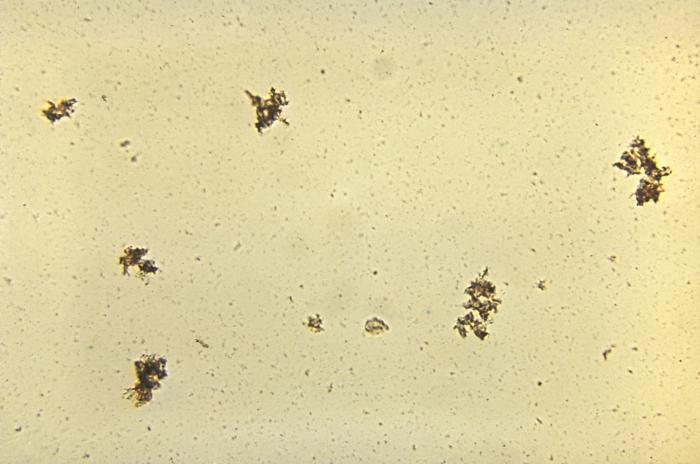
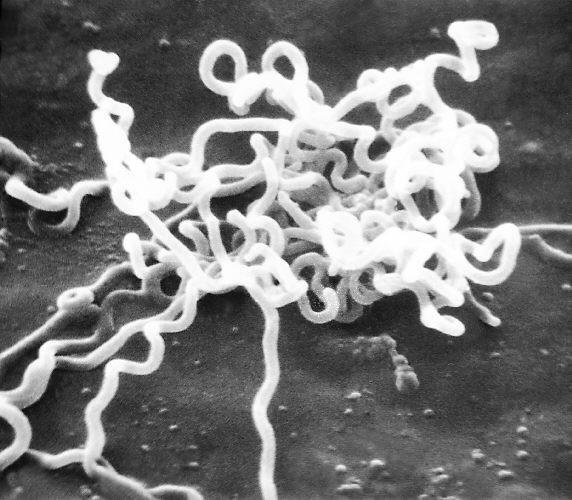
Treponema pallidumOverview: Treponema pallidum is a spirochetal bacterium often found growing in clusters (Figure 1). This pathogen has a central protoplasmic cylinder bound by a cytoplasmic membrane, a thin layer of peptidoglycan, and an outer membrane devoid of transmembrane proteins (Salazar et al., 2002) (Figure 2). There are three other structurally, serologically, and morphologically identical species belonging to the Treponema genus, namely, T. endemicum, T. pertenue, and T. carateum. These species can only be distinguished by genetic analysis. T. pallidum spiral shape, rapid rotation about its axis, and two to three flagella at each end allow a corkscrew like motility (Figure 3). The bacterium is approximately 6 to 20 μm (micrometres) in length and only 0.10 to 0.18 μm in width. It is microaerophilic and glucose-dependent; without glucose, oxygen impairs motility. T. pallidum contains one of the smallest prokaryotic genomes consisting of about 1000 kilobase pairs, and is responsible for the sexually transmitted disease, syphilis. Figure 1. An unstained micrograph depicting the growth of Treponema pallidum in small clusters.
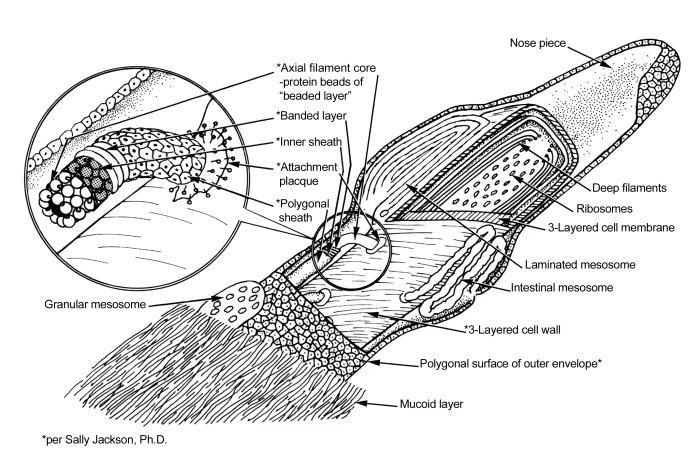
Figure 2. A schematic illustration of one end of a Treponema pallidum bacterium.
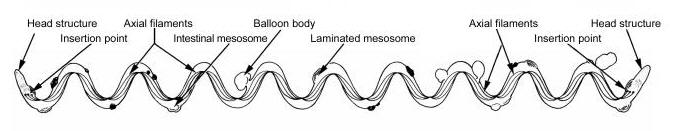
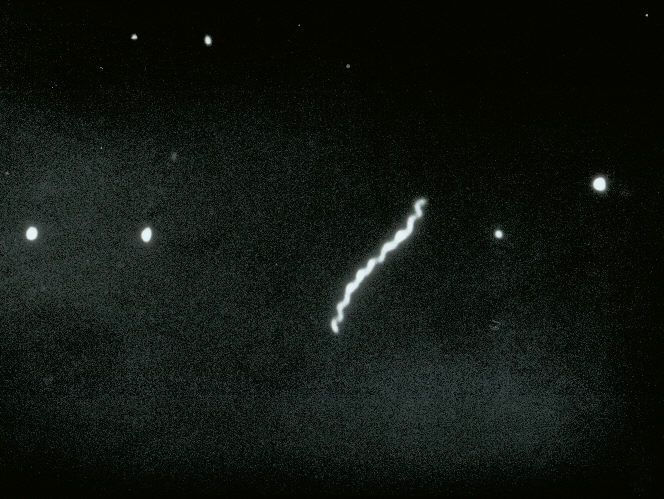
Figure 3. An anatomical illustration of a Treponema pallidum bacterium from a lateral perspective. From a lateral view, one is able to identify the cellular components along the length of a T. pallidum bacterium, which give this spiral bacterium (spirochete) a unique cellular morphology, and mode of motility. Identification: T. pallidum cannot be identified using Gram staining because it is too thin and it cannot be cultured in vivo, making it difficult to study in the lab. It may instead be characterized by several different methods, including the immunohistochemical Warthin-Starry-stain (WS-stain), electron microscopy, dark-field microscopy (Figure 4), immunofluorescence, or immunoperoxidase techniques with polyclonal antibodies. A positive darkfield result is an almost certain diagnosis of primary, secondary, or early congenital syphilis from suspected lesions or regional lymph nodes. In primary syphilis, the darkfield examination may provide a means by which to identify the etiologic agent of syphilis and diagnose the disease before antibodies to T. pallidum can be detected. The WS-stain shows a positive result for T. pallidum presence when it stains a dark brown colour, however this silver-staining technique presents problems with false-positives and false-negatives (Martín-Ezquerra et al., 2009). Figure 4. Using a darkfield microscopy technique, this photomicrograph reveals the presence of Treponema pallidum spirochetes. Darkfield microscopy involves the application of light rays in an oblique manner to microscopic specimens in order to illuminate these organisms, which are normally difficult to see using normal lighting techniques. Clinical Infections: T. pallidum is an obligate internal human parasite that is not native to the normal microflora and has no known non-human reservoir. Each subspecies causes a different infection; for instance, T. pallidum causes venereal syphilis, T. pertenue causes yaws, T. endemicum causes endemic syphilis (bejel), and T. carateum causes pinta (Antal et al., 2002). T. pallidum causes infections that may be divided into early stages (including primary and secondary lesions) and late stages. Early stage lesions are considered infectious and may last up to five years after the time of infection. There may be periods of latency between symptomatic episodes and before the late stage infection. Pinta only involves cutaneous lesions, while yaws and endemic syphilis also affect the mucous membranes and bones. Venereal syphilis is capable of affecting most organs and can cross the placenta (Antal et al., 2002). T. pallidum is distributed differently, depending on the stage of infection and the infecting species. In primary syphilis infections, T. pallidum is found in large numbers in the dermis and surrounding the vascular walls in a vasculotropic patterns. In secondary syphilis infection, this germ is distributed mainly in the lower layers of the epidermis in an epitheliotropic pattern (Martín-Ezquerra et al., 2009) (Figure 5). Venereal syphilis is characterized by genital ulcerations, skin rashes and in later stages, invasion of the central nervous system. The other three species of this genus commonly cause primary lesions and disseminated skin or mucous membrane lesions (Antal et al., 2002). Figure 5. An electron micrograph of Treponema pallidum on cultures of cotton-tail rabbit epithelium cells. Transmission:
Transmission of infection varies between the
diseases. Yaws is transmitted by
direct skin contact from a lesion and its
clinical expression varies according to
climatic conditions. Endemic syphilis is
transmitted via direct skin contact
with a lesion or by mucous membrane contact.
Pinta does not have a known
transmission; however, it is suspected to be
direct contact with a lesion.
Venereal syphilis is transmitted via intimate
contact. T. pallidum has been found to be a synergistic contributor to the risk of human immunodeficiency virus (HIV) infection. Since both HIV and syphilis are sexually transmitted infections, coinfection is common between them. Ulcers caused by syphilis provide an ideal entry point for the HIV virus and T. pallidum genetically alters macrophages to make the individual more susceptible to HIV (Karp et al., 2009). T. pallidum reproduces rapidly and can penetrate interjunctionally between vascular endothelia to gain access to various tissues. It is also poorly antigenic due to the fact that possible pathogenic antigens are hydrophilic and tethered to the inner cytoplasmic membrane, making them inaccessible to antibodies (Salazar et al., 2002). Some of the transmembrane proteins have been shown to be porins to destroy host-cells. Antigenic variation has also been suggested as a mechanism for persistence and prolonged host infection. Treatment: Infections caused by this species can be treated with various antibiotics, with penicillin being the most widely used. Ideally, these infections could be eradicated because they are not found in any other animal reservoirs, however poverty and illiteracy on hygiene make this difficult. Parenteral penicillin G is the only therapy with documented effect during pregnancy. For early syphilis, one dose of penicillin is sufficient. Those who are allergic to penicillin can effectively be treated with oral tetracycline or doxycycline; however, data to support this is limited. Ceftriaxone may be considered as an alternative therapy, although the optimal dose is not yet defined. However, cross-reactions in penicillin-allergic patients with cephalosporins such as ceftriaxone are possible. Azithromycin was suggested as an alternative. However, there have been reports of treatment failure due to resistance in some areas. References: Antal, G.M., Lukehart, S.A., & Meheus, A.Z. (1981). The endemic treponematoses. Microbes and Infection, 4: 83-94. Karp, G., Schlaeffer, F., Jotkowitz, A. & Riesenberg, K. (2009). Syphilis and HIV co-infection. European Journal of Internal Medicine, 20: 9-13. Martín-Ezquerra, G., Fernandez-Casado, A., Barco, D., Jucglà, A., Juanpere-Rodero, N., Manresa, J.M., Soares de Almeida, L.M., & Rodríguez-Peralto, J.L. (2009). Treponema pallidum distribution patterns in mucocutaneous lesions of primary and secondary syphilis: an immunohistochemical and ultrstructural study. Human Pathology, 40; 624-630. Salazar, J.C., Hazlett, K.R.O., & Radolf, J.D. (2002). The immune response to infection with Treponema pallidum, the stealth pathogen. Microbes and Infection, 4: 1133-1140. |